イメージングで迫るリアルタイムのオートファジー像
オートファジーは細胞内の分解機構の1つであり、最近、研究の進歩が著しい分野である。オートファジー研究は出芽酵母に続き哺乳類でも因子群の同定が進み、その分子メカニズムが徐々に明らかになってきた。その草分け的存在であり、オートファジー研究の牽引者の1人である東京医科歯科大学の水島昇氏と、2010年より水島研究室でオートファジーのライブイメージングで研究を進める本田郁子氏の話からオートファジー研究の新しい一面が見えてきた。

水島昇氏 (みずしま のぼる)
1996年東京医科歯科大学大学院医学研究科修了。博士(医学)
日本学術振興会特別研究員、さきがけ研究21研究員、基礎生物学研究所助手、東京都臨床医学総合研究所室長を経て、2006年より東京医科歯科大学大学院医歯学総合研究科細胞生理学分野教授。

本田(小山)郁子氏 (ほんだ いくこ)
2002年名古屋大学大学院理学研究科生命理学専攻博士課程後期満了。2005年名古屋大学理学研究科にて博士号(理学)取得。
ERATO楠見膜組織能プロジェクト研究員、ICORP膜機構プロジェクト研究員を経て、2010年より日本学術振興会特別研究員(RPD)。
新陳代謝への興味から始まったオートファジー研究
「もともと学生時代から新陳代謝に関心を持っていました。当時、すでに転写などの生体内での合成についてはよく分かっていましたが、タンパク質や細胞がどのように無くなるのかといったことは全くわかっていなくて、ずっと興味を持っていました」と語る水島氏がオートファジーの研究に取組み始めたのは、博士研究員時代から。もともと医学部で免疫系の研究に取組んでいたが、基礎生物学研究所の大隅良典氏(現・東京工業大学特任教授)がオートファジーに関わる研究を行っていることを知り、卒業後にその世界に飛び込むことを決めた。
研究を始めた1996年当時、大隅研究室では出芽酵母を使ったオートファジーの遺伝学的な解析による因子の同定が盛んに行われていた。そこで携わったのが、Atg12(当時の名称はApg12)だった。この研究の中でAtg5(当時の名称はApg5)とAtg12が共有結合体を形成しているという知見にたどり着くことになる※1。Atg12、Atg5のホモログが哺乳類にも存在することがわかり、同じように共有結合体を作ることも確認できた※2。分子レベルで酵母と高等真核生物の研究が結びつき、今につながる高等真核生物でのオートファジー研究が幕を開けた。
強力な観察ツールの確立
「オートファジー研究を進めるうえで一番大きかったのは、オートファジーを蛍光顕微鏡で観察できるトランスジェニックマウスを作ることができたことですね。それまでの研究では、オートファジーがどこで起こっているかを調べるためには、切片を作って電子顕微鏡で観察する必要があったのですが、このマウスのおかげで、いつ、どこで起こるのかということがわかるようになりました。これがなかったら、受精卵でオートファジーが起こっているということも、わからなかったと思います」。この方法を初めて用いた2004年の論文では、マウスのオートファジー因子LC3とGFPの融合タンパク質を使い、肝臓、筋肉、膵外分泌細胞、糸球体、胸腺皮質など、あらゆる部位でのオートファジーが鮮やかに浮かび上がっている※3。この頃からオリンパスの倒立顕微鏡IXシリーズとDSU(ディスク走査型顕微鏡)を使っていますね、とオートファジーのイメージングを振り返る。
リアルタイムで見えてきた因子群間の関係
「日本にいるなら超一流の研究室に行きなさいという恩師のアドバイスを参考にして、自分が興味を持っている膜と細胞に関わる研究のできるところで、世界トップを行く研究室を探して水島研を選びました」。こう語る本田氏は、膜関連因子のイメージングに10年以上携わってきたという経歴の持ち主だ。水島研ではこれまでに個体を使ったオートファジーのイメージングと並行して、細胞を使ったライブイメージングも行ってきた。本田氏が2010年に新たなメンバーとして加わったことで、後者の研究が加速している。
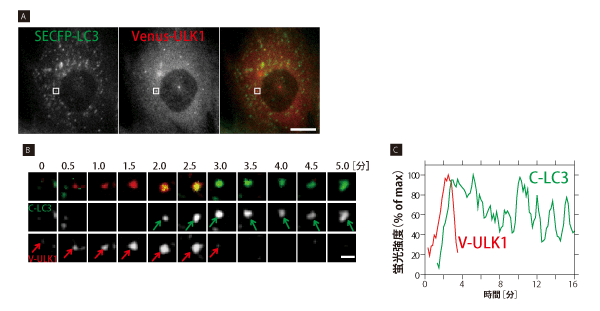
オートファジーの因子は大きく5つのグループに分かれる(表1)。遺伝学的な解析から、オートファジーの一連のカスケードにおけるこれらのグループの関係は明らかにされていた。しかし、リアルタイムで時系列変化を追った研究はこれまで行われてこなかった。「本田さんがいなかったらこれだけ根気のいる仕事は進まなかったと思いますね」という水島氏の言葉に現れるように、本田氏の成果の蓄積がリアルタイムのオートファジーの進行をとらえ始めている。例えば、オートファゴソーム因子であるLC3にSECFPを、オートファゴソーム前駆体因子であるULK1にVenusを融合したタンパク質を発現する細胞でライブイメージングを行うと、始めにVenus-ULK1のドットが現れ、次いで共局在する形でSECFP-LC3のドットが現れると、次第にVenus-ULK1のドットがフェードアウトしていく様子が見事に観察されている(図1)※4。このように各グループに属する代表的な因子に異なる蛍光標識し、ライブイメージングで観察したことで、遺伝学では明らかにできなかったオートファジーのカスケードが姿を現してきたのだ。
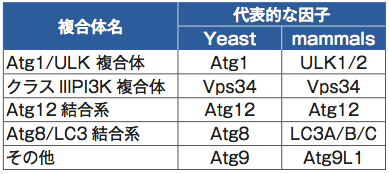
表 1 オートファゴソーム因子のグループと代表的な因子
哺乳類の代表的な因子とそれに対応する出芽酵母の因子を記した。
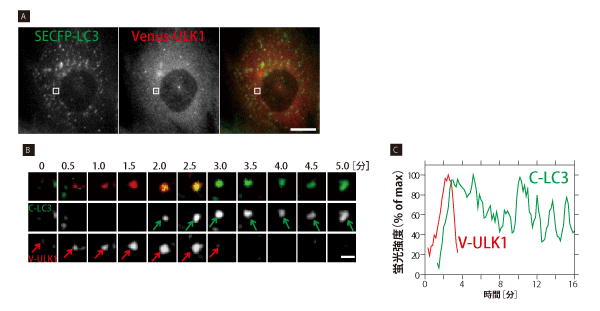
図 1 オートファゴソーム因子 LC3、ULK1 の局在変化
(A) SECFP-LC3(緑), Venus-ULK1(赤)発現MEF細胞を、アミノ酸、血清飢餓条件でオートファジー誘導約2時間後、2 色ライブ観察を行った。10 秒毎に約 20 分間録画した。図は動画中の1フレーム画像を示す。ULK1は栄養シグナルを感知で きる上流Atg因子、LC3はオートファゴソームに局在する下流Atg因子である。定常状態では、SECFP-LC3, Venus-ULK1 はほとんどが細胞質中に拡散しているが、オートファジー誘導条件では図のように細胞内にドット状に蓄積し、オートファゴソー ムを形成する。倒立顕微鏡IX81, PlanApo 60x NA=1.42(共にオリンパス), CoolSNAP HQ2(日本ローパー)使用。スケー ルバーは20μm。
(B) オートファゴソーム形成過程の一例 (Aの枠部分)。緑 と 赤 矢印はSECFP-LC3, Venus-ULK1の同一ドットを示す。 始めにVenus-ULK1(赤)のドット状構造が現れ、約 3分で蓄積極大に達した後、急速に消失した。SECFP-LC3(緑)ドット はVenus-ULK1上 に遅れて現れ、観察時間内に消失しなかった。スケールバーは2mm。
(C) Bで示したSECFP-LC3(緑), Venus-ULK1(赤)ドット状構造の蛍光強度-時間変化。
「将来の目標はオートファジーの膜脂質のソースがどこなのか、膜がどのように閉じるのかといった仕組みにイメージングで迫ることです」と語る本田氏。現在メインで使用する倒立顕微鏡IX81に加え、新たに導入した共焦点レーザー走査型顕微鏡FV1000-Dの高分解能の像から、また新しいオートファジーのメカニズムが見えて来ることも期待される。ここでの発見は、マウスの個体中での制御や何のためにオートファジーが起こるのかといった水島氏がこれからの課題とする問題の本質的な理解に通じるはずだ。

共焦点レーザ走査型顕微鏡FV1000-D(倒立型電動リサーチ顕微鏡IX81-ZDC仕様)
※1 Mizushima, N. et al. (199 8). A protein conjugation system essential for autophagy. Nature, 395, 395-398.
※2 Mizushima, N. et al. (1 998). A new protein conjugation system in human. The counterpart of the yeast Apg12p con jugation system essential for autophagy. J. Biol. Chem. 273, 33889-33892.
※3 Mizushima, N. et al. (2 004). In vivo analysis of autophagy in response to nutrient starvation using transgenic mice expressing a fluorescent autophagosome marker. Mol. Biol. Cell , 15, 1101- 1111.
※4 2011年第34回日本分子生物学会年会にて発表。
オリンパス株式会社
[所在地]本社:東京都新宿区西新宿2-3-1新宿モノリス
[TEL]03-6901-4456(ライフサイエンス事業本部)
[URL]http://www.olympus.co.jp/jp/lisg/bio-micro/
●本記事に関するお問い合わせ
TEL03-6901-4456(ライフサイエンス事業本部)